وظیفه آنها : انتقال (ترارسانی) دادههای عصبی : انجام از راه هدایت تکانههای الکتریکی
این سلول ها در بیشتر موارد تقسیم نمیشوند.
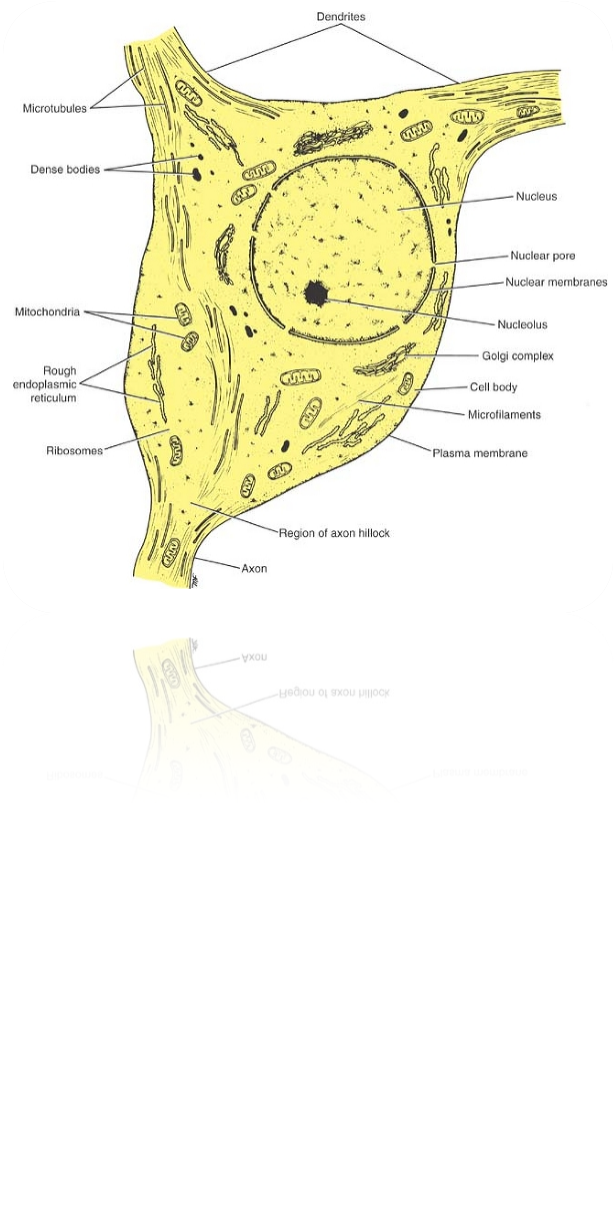
جسم سلولی : بخش هستهدار سلول عصبی، جسم سلولی یا soma خوانده میشود. شکل و اندازه جسم سلولی در نورونهای مختلف، متفاوت است. بسیاری از آنها به شکلهای مدور، بیضوی، هرمی، ذوزنقهای و ستارهای هستند و قطر آنها از چند میکرون تا چند صد میکرون متفاوت است. جسم سلولی دارای هسته مشخص و سیتوپلاسم ویژهای است که آن را پریکاریون (Perikaryon) مینامند.
نوروپلاسم : سیتوپلاسم سلول عصبی را نوروپلاسم (Neuroplasm) مینامند. در نوروپلاسم، اندامکهای ریزی وجود دارد که در سایر سلولهای زنده بدن نیز یافت میشود. از جمله این اندامکها عبارتند از:
1)شبکه آندوپلاسمی یا درون سیتوپلاسمی با دانههای ریبوزوم که بر روی آن قرار دارند: در پروتئینسازی و نقل و انتقال آنها نقش اساسی داشته و در برخی نواحی، دانههای درشتی را میسازند که رنگهای قلیایی را به خود جذب میکنند که اجسام نیسل(Nissl)خوانده میشوند.
2)میتوکندریها: انرژی لازم را برای زندگی سلول فراهم میکنند.
3)دانههای لیزوزوم: این دانهها محتوی آنزیمهایی هستند که میتوانند ذرات خارجی و مواد زاید را تجزیه و تخریب کنند.
4)دستگاه گلژی: در غشاسازی و بستهبندی ترشحات نورون نقش اساسی دارد.
5)نوروفیبریلها و نوروتوبولها: نوروفیبریلها همان رشتههای بسیار نازک و نوروتوبولها هم، لولههای بسیار نازکی هستند که در نوروپلاسم وجود دارند. این رشتهها و لولهها نوعی اسکلت سیتوپلاسمی تشکیل میدهند که به حفظ شکل نورون و تار عصبی کمک میکنند. این لولهها و رشتهها توسط میکروسکوپهای الکترونیکی قویتری نسبت به میکروسکوپهایی که توانایی مشاهده اندامکها و سلول عصبی را دارد، دیده میشوند
هسته : هسته نورون نیز تفاوتی با هسته سایر سلولها ندارد. در درون آن شیره هسته یا نوکلئوپلاسم، یک یا چند هستک(نوکلئول) و تودههای کروماتین دیده میشود
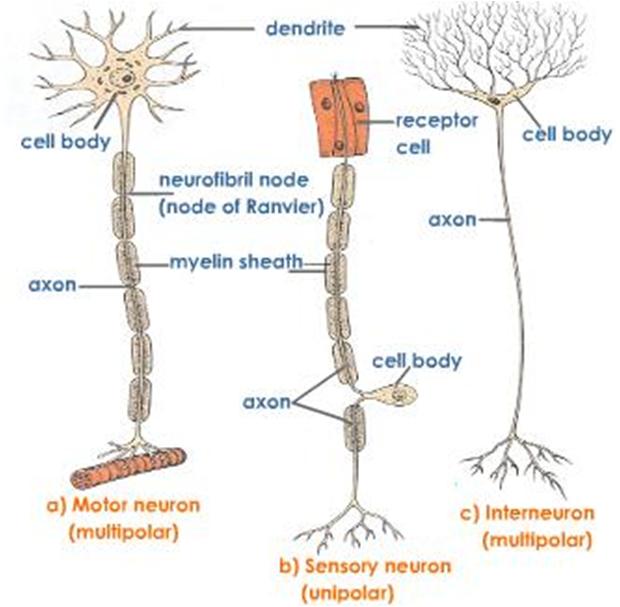
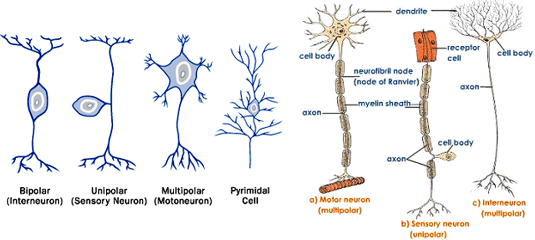
نورونها از نظر طرز خارج شدن تارهای عصبی از جسم سلولی به سه گروه یکقطبی، دوقطبی و چندقطبی تقسیم میشوند:
نورونهای یکقطبی: در این نورونها آکسون و دندریتها از یک ناحیه جسم سلولی خارج میشوند. نمونه این نورنها در عقدههای نخاعی انسان دیده میشود که گیرندههای حسهای پیکری هستند. اغلب نورونهای بیمهرگان نیز یکقطبی هستند.
نورونهای دوقطبی: در این نورونها، آکسون از یک ناحیه و دندریتها از ناحیه دیگر جسم سلولی بیرون میآیند. گیرندههای بویایی که در مخاط زرد بینی قرار دارند و برخی نورونهای شبکیه چشم از این نوع هستند.
نورونهای چندقطبی: تعداد این نورونها از دو نوع قبلی بیشتر است. در این نورونها آکسون از یک ناحیه و دندریتها از چند ناحیه دیگر جسم سلولی خارج میشوند. در این نورونها تشخیص آکسون از دندریتها بسیار ساده است. نورونهای حرکتی شاخ پیشین نخاخ، نورونهای پورکنژ مخچه و نورونهای هرمی شکل قشر مخ همگی از نوع چندقطبی ولی با شکلهای متفاوت هستند.
در بعضی قسمتهای دستگاه عصبی، نورونهایی که فاقد آکسون است شناسایی شده است که این نورونها فقط قادرند تحریک عصبی را به نورونهای مجاور خود منتقل کنند.
غلاف میلین یک لایه لیپوپروتئینی است که بر روی بسیاری از دندریت های بلند و آکسون ها، تشکیل میشود. نقش اصلی پوشش میلین، ایجاد نارسانایی بیشتر بر روی سطح تار عصبیها است که این امر باعث افزایش سرعت هدایت پیامهای الکتریکی در درازای تار میشود. میلین افزون بر افزایش سرعت انتقال پیامهای عصبی در طول رشتههای عصبی، وظیفه نگهداری از سلولهای عصبی را نیز به عهده دارد.
همه طول تار از میلین پوشیده نشدهاست. بخشهایی که پوشش میلین وجود ندارد گره رانویه نامیده میشوند. در رشتههای میلیندار پیام عصبی بسیار تندتر منتقل میشود.
از دیدگاه میلینه بودن، نورون ها بر ۲ گونه هستند:
نورون های بدون میلین: سرعت پایینی در انتقال پیامهای عصبی، ایجاد بخش خاکستری مغز و نخاع
نورون های میلین دار: سرعت انتقال در این ها بیش از ۱۰۰ برابر نورورن های بی میلین، ساختن بخش سفید مغز و نخاع
میلین توسط سلولهای الیگودندروسیت (در سیستم عصبی مرکزی) و سلولهای شوان (در سیستم عصبی محیطی به دور تارهای عصبی ساخته میشود. سلولهای نوروگلیا عمل تغذیه و از بین بردن میکروبها را هم انجام میدهند.
از بین رفتن پوشش میلین موجب نارسایی در هدایت پیامهای عصبی و بنابراین بروز بیماریهای عصبی همچون اسکلروز چندگانه یا همان ام. اس میشود. دانشمندان سالها است برای یافتن شیوهای که با آن بتوان بدن را به بازسازی دوباره میلین واداشت، پژوهش میکنند.
پتانسیل عمل بر روی غشای تحریک پذیر یک سلول عصبی پدید میآید، طول اکسون را طی میکند و وظیفه پیغام رسانی را بر عهده دارد.
وقتی نورون تحریک میشود. وضعیت بارهای الکتریکی در دو سوی غشای آن در نقطه تحریک تغییر میکند. یعنی سطح خارجی نقطه تحریک شده منفی و سطح داخلی آن مثبت میشود.
علت آن است که غشای نورون در نقطه تحریک شده ، نسبت به سدیم نفوذ پذیر میشود. با هجوم سدیم به درون نورون سطح بیرونی منفی و سطح داخلی مثبت میشود. تغییر بار الکتریکی در نقطه تحریک شده باقی نمیماند و نقطه به نقطه در طول تار عصبی حرکت میکند و جریان یا پیام عصبی را پدید میآورد. بار الکتریکی هر نقطه پس از تحریک فوری به حال اول بر میگردد.
پتانسیل الکتریکی نورون در هنگام تحریک آن پتانسیل عمل نام دارد.
قبل از توضیح کانال و پمپ سدیم و پتاسیم باید مفهوم شیب غلظت یونی را معین کنیم. غلظت پتاسیم درون سلول زیادتر از بیرون سلول است و در مورد سدیم دقیقاٌ برعکس است. بنابراین جهت شیب غلظت پتاسیم از داخل به خارج (غلظت زیادتر به کمتر) است و شیب غلظت سدیم از خارج به داخل است. به این ترتیب پتاسیم تمایل دارد در جهت شیب غلظت خود حرکت کند یعنی از تراکم بالا در درون سلول به تراکم کم در خارج سلول برود و جهت شیب غلظت سدیم هم برعکس پتاسیم است.
نکته : شیب غلظت سدیم از خارج سلول به داخل و شیب غلظت پتاسیم از داخل سلول به خارج است.
پتانسیل آرامش :
زمانی که نورون در حال استراحت است و توسط محرک و پیام عصبی ناشی از آن تحریک نمیشود، داخل غشای نورون، پتانسیل منفی تر از خارج نورون و برابر 65- میلی ولت میباشد. این پتانسیل منفی داخل نورون توسط شیب غلظت سدیم و پتاسیم و پمپ سدیم ـ پتاسیم ایجاد میشود.
در حالت استراحت پتاسیم در جهت شیب غلظت خود از سلول خارج میشود.(چون نفوذپذیری غشا به پتاسیم زیاد است) ولی سدیم به همین شکل نمیتواند در جهت شیب غلظت خود وارد سلول شود چون نورون نسبت به سدیم خیلی نفوذپذیر نیست. لذا علی رغم حرکت یونها در جهت شیب غلظتشان، به دلیل یکسان نبودن نفوذپذیری غشا نسبت به یونها، در دو سوی غشای نورون یونهای پتاسیم بیش تر خارج میشوند و به همان نسبت یون سدیم وارد نمیشود.
داخل غشا بار مثبت زیادتری از دست میدهد . و پتانسیل منفی تری نسبت به خارج غشا پیدامیکند و این بار منفی به خاطر کمبود یون مثبت است.
پتانسیل عمل: زمانی که پیام عصبی به منطقه ای از نورون میرسد که در حال آرامش است، تعادل بار را در دو سوی غشای این ناحیه به هم میزند که در نتیجهی آن داخل غشا مثبت و خارج آن منفی میشود. کانالهای سدیم و پتاسیم در ایجاد پتانسیل عمل دخیل هستند.( کانال سدیم در شروع پتانسیل عمل و کانال پتاسیم در ادامهی پتانسیل عمل نقش دارند). مراحل آن :
1) ابتدا با رسیدن پیام عصبی، کانالهای سدیم که به ولتاژ پیام عصبی حساس هستند باز میشوند. یونهای سدیم که تا حالا پشت در،مانده بودند و غشا برایشان ناز میکرد و نفوذپذیری نشان نمیداد! به داخل نورون هجوم میآورند که به تبع آن پتانسیل داخل غشا سریعاً مثبت میشود و خارج غشا که از بار مثبت (یون سدیم) خالی شده بار منفی پیدا میکند.
2) با اختلاف زمان خیلی خیلی ناچیز از باز شدن کانالهای سدیم، کانالهای پتاسیم باز میشوند و یونهای پتاسیم درون سلول که تا حالا به آرامیدر جهت شیب غلظت خود خارج میشدند، با سرعت به بیرون میروند. خروج پتاسیم باعث جبران بار مثبت از دست رفتهی خارج نورون میشود و پتانسیل دو سوی غشا تقریباً به حالت آرامش برمیگردد.
چون کانالهای پتاسیم تقریباً بلافاصله بعد از کانالهای سدیم باز میشوند، پتانسیل عمل در چند لحظهی بسیار کوتاه بیشتر برقرار نمیماند. اما در عین حال هنوز غشاء تعادل یونی ( تعادل سدیم و پتاسیم در دو سوی غشاء) را به دست نیاورده و این کار توسط پمپ سدیم ـ پتاسیم انجام میشود.
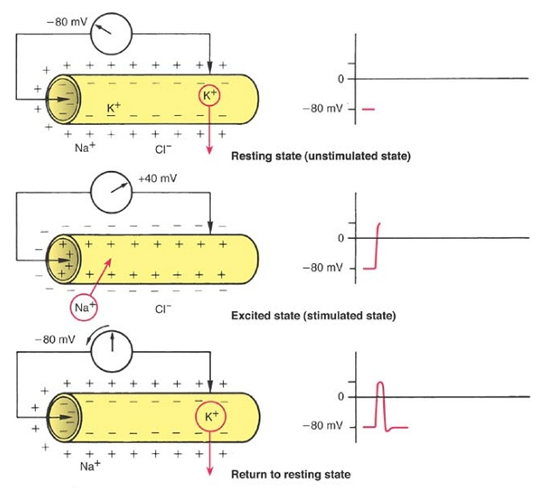


از شروع پتانسیل عمل تا بازگشت به حالت آرامش به ترتیب 3 مرحله وجود دارد:
1) باز شدن کانالهای سدیم 2) باز شدن کانالهای پتاسیم و بسته شدن کانالهای سدیم 3) بسته شدن کانالهای پتاسیم و شروع فعالیت پمپ سدیم- پتاسیم.
مرحلهی1 دپلاریزاسیون و مراحل 2 و 3 را رپلاریزاسیون مینامند. پنانسیل عمل شامل مراحل 2و 1 و پتانسیل استراحت شامل مراحله 3 میباشد.
پس از پتانسیل عمل غشا نسبت به هر دو یون سدیم و پتاسیم نفوذناپذیر میشود . در این مرحله یونهای سدیم و پتاسیم توسط کانالها منتقل نمیشوند و تبادل یونها تنها توسط پمپ سدیم- پتاسیم انجام میگیرد.
غشاء در تمامی مراحل پتانسیل عمل و آرامش نسبت به پتاسیم نفوذپذیر است و نسبت به سدیم نفوذپذیری بسیار ناچیزتری دارد. اصولا نشت و عبور یون از غشا بسیار کم انجام میشود؛ به جززمان بعد از پتانسیل عمل که غشا کلاً به یونها نفوذناپذیر است، (یونها فقط از طریق نشت ازعرض غشاء رد میشوند نه از طریق کانال)
تنها در زمان پتانسیل عمل یعنی زمانی که کانالهای سدیم باز میشوند غشا به سدیم .اجازه ورود در مقادیر بالا را می دهد و نفوذناپذیریاش به سدیم خیلی زیاد میشود.
پتانسیل آرامش (داخل غشا منفی و بیرون آن مثبت) در تمام سلولها، چه عصبی چه غیرعصبی وجود دارد ولی پتانسیل عمل که نشانهی تحریک پذیری است ویژگی سلولهای عصبی است و در همه سلولها دیده نمیشود.
منظور از شیب غلظت تفاوت غلظت و تراکم است و معمولاٌ از تراکم بیشتر به کمتر در نظر گرفته میشود. مثلاً «شیب غلظت سدیم از خارج به داخل است» یعنی غلظت سدیم در خارج بیش تر است و در داخل کمتر.
در منحنی پتانسیل غشا، بخش بالا روی منحنی که معرف باز شدن کانالهای سدیمیاست، مرحلهای از پتانسیل عمل (دپلاریزاسیون) را نشان میدهد و بخش پایین رو منحنی که معرف بازشدن کانالهای پتاسیمیاست مرحله بازگشت به حالت اولیه (رپلاریزاسیون) را نشان میدهد.
نحوهی انتقال سدیم و پتاسیمیکه توسط پمپ در زمان استراحت جابجا میشوند چون با مصرف انرژی (ATP) و در خلاف شیب غلظتشان میباشد، انتقال فعال نامیده میشود. ولی نحوهی انتقال سدیم و پتاسیم انتقالی در زمان پتانسیل عمل چون توسط کانال و بدون مصرف انرژی و در جهت شیب غلظتشان حرکت میکنند انتشار میباشد.
ویژگیهای پتانسیل آرامش: 1) کانالهای سدیم بسته و کانالهای پتاسیم باز هستند. 2) پمپ سدیم ـ پتاسیم فعال است. 3) غشا به سدیم نفوذپذیری کمتری از پتاسیم دارد.
پایان پتانسیل عمل زمانی است که کانال پتاسیم بسته میشود .
پمپ سدیم ـ پتاسیم در غشای همهی سلولها وجود دارد و باعث ایجاد پتانسیل آرامش در تمام سلولها میشود.
تحریک پذیری
تحریک پذیری از ویژگیهای هر سلول زنده است، اما نورونها این خاصیت را بهتر نشان میدهند . به عاملی که باعث تحریک میشود محرک میگویند. محرک ممکن است الکتریسته ، تغییر دما ، نور ، مواد شیمیایی ، ضربه، فشار و یا صدا باشد. برای آنکه محرکی بتواند نورون را تحریک کند. نباید شدت آن از حد معینی کمتر باشد. این حد معین از شدت تحریک را شدت آستانه میگویند.
هدایت و انتقال پیام عصبی
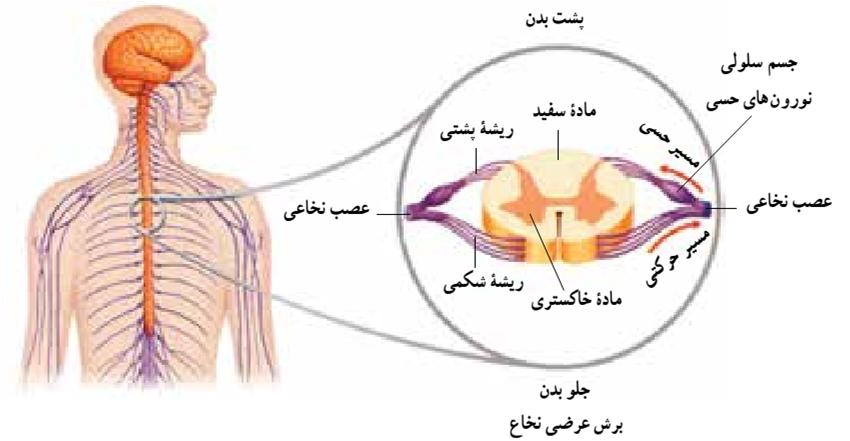
محل ارتباط دو نورون ، یا نورون و سلول ماهیچهای را سیناپس می گویند. در محل سیناپسها ، پایانههای آکسون به دندریتها ، به جسم سلولی نورون دیگر و یا به سلول ماهیچهای نچسبدهاند بلکه فضای کوچکی به نام فضای سیناپسی در میان آنها وجود دارد. منظور از انتقال پیام عصبی ، انتقال پیام در محل سیناپسهاست. انتقال پیام عصبی با واسطه مواد شیمیایی مخصوص صورت میگیرد.
همایه یا سیناپس یک ساختار زیستی در پایانه آکسونها است که از راه آن یک سلول عصبی پیام خود را به دندریت یک نورون دیگر یا یاخته ماهیچهای یا یک غده میفرستد.
اگرچه تعداد بسیار زیادی سیناپس در مغز انسان وجود دارد، آنها می توانند در دو نوع دسته بندی شوند: الکتریکی و شیمیایی